A new paper makes a strong case that the Huns, a group of "barbarians" (in the eyes of Roman historians) who made multiple attempts to invade the Roman empire, spoke a Paleo-Siberian language (to which the Na-Dene languages of North America, such as Navajo, are distantly related), rather than a Turkic language, as conventional wisdom in historical linguistics prior to this paper had wrongly believed.

The Xiōng-nú were a tribal confederation who dominated Inner Asia from the third century BC to the second century AD. Xiōng-nú descendants later constituted the ethnic core of the European Huns. It has been argued that the Xiōng-nú spoke an Iranian, Turkic, Mongolic or Yeniseian language, but the linguistic affiliation of the Xiōng-nú and the Huns is still debated.
Here, we show that linguistic evidence from four independent domains does indeed suggest that the Xiōng-nú and the Huns spoke the same Paleo-Siberian language and that this was an early form of Arin, a member of the Yeniseian language family. This identification augments and confirms genetic and archaeological studies and inspires new interdisciplinary research on Eurasian population history.
New linguistic findings show that the European Huns had Paleo-Siberian ancestors and do not, as previously assumed, originate from Turkic-speaking groups. The joint study was conducted by Dr. Svenja Bonmann at the University of Cologne's Department of Linguistics and Dr. Simon Fries at the Faculty of Classics and the Faculty of Linguistics, Philology and Phonetics at the University of Oxford.The results of the research, "Linguistic evidence suggests that Xiōng-nú and Huns spoke the same Paleo-Siberian language," have been published in the journal Transactions of the Philological Society.On the basis of various linguistic sources, the researchers reconstructed that the ethnic core of the Huns—including Attila and his European ruling dynasty—and their Asian ancestors, the so-called Xiongnu, shared a common language. This language belongs to the Yeniseian language family, a subgroup of the so-called Paleo-Siberian languages. These languages were spoken in Siberia before the invasion of Uralic, Turkic and Tungusic ethnic groups. Even today, small groups who speak a Yeniseian language still reside along the banks of the Yenisei River in Russia.From the 3rd century BCE to the 2nd century CE, the Xiongnu formed a loose tribal confederation in Inner Asia. A few years ago, during archaeological excavations in Mongolia, a city was discovered that is believed to be Long Cheng, the capital of the Xiongnu empire. The Huns, in turn, established a relatively short-lived but influential multi-ethnic empire in southeastern Europe from the 4th to 5th centuries CE.Research has shown that they came from Inner Asia, but their ethnic and linguistic origins have been disputed until now, as no written documents in their own language have survived. A great deal of what we know about the Huns and the Xiongnu is therefore based on written documents about them in other languages; for example, the term "Xiōng-nú' derives from Chinese.

[Based on the "World Topographic Map" by Esri. Sources: Esri, HERE, Garmin, Intermap, INCREMENT P, GEBCO, USGS, FAO, NPS, NRCAN, GeoBase, IGN, Kadaster NL, Ordnance Survey, Esri Japan, METI, Esri China (Hong Kong), OpenStreetMap contributors, GIS User Community, Simon Fries. Created with QGIS 3.36.]. Credit: Transactions of the Philological Society (2025). DOI: 10.1111/1467-968X.12321From the 7th century CE, Turkic peoples expanded westwards. It was therefore assumed that the Xiongnu and the ethnic core of the Huns, whose own westward expansion dates back to the 4th century CE, also spoke a Turkic language. However, Bonmann and Fries have found various linguistic indications that these groups spoke an early form of Arin, a Yeniseian language, in Inner Asia around the turn of the millennium."This was long before the Turkic peoples migrated to Inner Asia and even before the splitting of Old Turkic into several daughter languages. This ancient Arin language even influenced the early Turkic languages and enjoyed a certain prestige in Inner Asia. This implies that Old Arin was probably the native language of the Xiongnu ruling dynasty," says Bonmann.Bonmann and Fries analyzed linguistic data based on loan words, glosses in Chinese texts, proper names of the Hun dynasty as well as place and water names. Taken by itself, the data on each of these aspects would have comparatively little significance, but taken together it is hard to argue with the conclusion that both the ruling dynasty of the Xiongnu and the ethnic core of the Huns spoke Old Arin.The findings of the study also made it possible for the first time to reconstruct how the Huns came to settle in Europe: For the two researchers, place and water names still prove today that an Arin-speaking population once left its mark on Inner Asia and migrated westwards from the Altai-Sayan region. Attila the Hun probably also bears an ancient Arin name: Until now, "Attila" was thought to be a Germanic nickname ("little father"), but according to the new study, "Attila" could also be interpreted as a Yeniseian epithet, which roughly translates as "swift-ish, quick-ish."The new linguistic findings support earlier genetic and archaeological findings that the European Huns are descendants of the Xiongnu. "Our study shows that alongside archaeology and genetics, comparative philology plays an essential role in the exploration of human history. We hope that our findings will inspire further research into the history of lesser-known languages and thereby contribute further to our understanding of the linguistic evolution of mankind," concludes Fries.
In the body text, a section of the paper explores the previous conventional wisdom and its difficulties:
Although direct evidence is lacking, Iranian, Turkic and Mongolic languages have all been proposed as the language of the ruling dynasty of the Xiōng-nú (cf. e.g. Shiratori 1900; Benzing 1959; Pritsak 1982; Bailey 1985; Dybo 2007; Janhunen 2010; Beckwith 2018; Beckwith 2022) and of the Huns (cf. e.g. Doerfer 1973; Pritsak 1982; Savelyev 2020; Savelyev & Jeong 2020), because in the 1st millennium AD languages from these three families were spoken in Inner Asia. Inscriptions dating between the 4th and 9th century AD demonstrate that Iranian languages (Sogdian, early 4th to 6th century AD, Sims-Williams 2011; Vovin 2018) and Mongolic ones (Khüis Tolgoi and Bugut inscriptions of the 5th–6th centuries AD, Vovin 2018) as well as, much later, Turkic languages (isolated Turkish phrases in Bactrian manuscripts of the 7th century AD, Orkhon and Yenisei Kirgiz inscriptions between the early 8th and 9th century AD, Erdal 2004: 4–8) were spoken in the territory between the Yenisei River in the West, the Tian Shan range in the South and Mongolia in the East. Other Indo-European languages were spoken in oasis cities along the northern and southern ridges of the Takla Makan desert in the 1st millennium AD including Indo-Iranian (Iranian Khotanese and Tumshuqese Saka, Bactrian, Indo-Aryan Prakrit, Sanskrit) and ‘Tocharian’ languages (Agnean and Kuchean).However, this linguistic situation of a coexistence of Iranian, Turkic and Mongolic in Inner Asia can only be reliably established as such for the late 1st millennium AD. Hypotheses on an Iranian, Mongolic or Turkic identity of the Xiōng-nú primarily rest on written sources post-dating the Xiōng-nú era.
While the theoretical possibility of a Mongolic or Turkic presence in Inner Asia already at the beginning of the common era cannot be ruled out a priori, it is important to note that there is, on the other hand, also no robust evidence – especially from textual sources – that could directly imply or prove a Turko-Mongolic presence in this area at such an early date.
The earliest sources from the Tarim Basin and the territories alongside the Oxus River/Amu Darya (Chorasmia, Sogdia, Bactria) only document Indo-European languages from the Indo-Iranian and ‘Tocharian’ branches (to which might be added, as a cultural import, also Ancient Greek in Macedonian colonies). Judging by more indirect evidence – especially loanwords in other languages, toponyms, etc. – other Iranian languages, namely different Sakan varieties (Tremblay 2005) and ‘Old Steppe Iranian’ (Bernard 2023), must have been spoken in the steppe corridor from the Kazakh steppe to Dzungaria, and perhaps even to Gansu (see Beckwith 2022). It is only centuries later, namely in the Migration Period of the 5th–6th centuries AD, that a (Para-)Mongolic language might be attested in Inner Asia (Vovin 2018), and fragments of this (Para-)Mongolic language, in turn, are still much earlier documented than the earliest secure Turkic words dating from the 7th century AD.There is thus neither direct nor indirect evidence supporting the claim of a Mongolic or Turkic presence in Inner Asia between the 3rd century BC and the 2nd century AD, and the hypothesis of a Mongolic or Turkic identity of the ethnic core of the Xiōng-nú (as proposed by Benzing 1959, Pritsak 1982; Tenišev 1997; Dybo 2007; Janhunen 2010; Savelyev 2020) is thus rather unlikely from the outset, as is the hypothesis of a completely unknown or unclassifiable language without any living descendants (as proposed by Doerfer 1973). The same applies to the Huns: there is a complete lack of evidence supporting claims of a Turkic presence among the Huns.1 On the other hand, an Iranian component in the Xiōng-nú Empire is possible, and indeed quite likely, although, as we intend to point out with the present study, such Indo-European ethnicity must not necessarily have been shared by the ruling dynasty or ethnic core of the Xiōng-nú (pace Bailey 1985; Beckwith 2022) or the Huns.Concerning such an Iranian component, (Beckwith 2018, 2022) has argued recently that Xiōng-nú words preserved in Chinese texts are indicative of an Iranian language, which he calls ‘East Scythian’. However, his interpretation depends on a reconstruction of the Old and Middle Chinese pronunciation of Chinese signs which significantly differs from established reconstructions such as the classic one of Pulleyblank, and which has also been criticised by Vovin et al. (2016: 129–30). In addition to this, his Iranian etymologies must be met with serious doubts. For instance, the ethnonym ‘Aryan’, which is amply attested in many Indo-Iranian languages, is given by Beckwith with a word-initial laryngeal sound (discussion in Beckwith 2022: 183–86, cf. particularly p. 186): ‘East Scythian *ḥarya [ɣa.rya] “noble, royal; Scythian” → Old Chinese *ḥaryá 夏/*ḥâryá 華 “royal; Chinese, China”’. This would indeed be a remarkable Iranian word form, because no Indo-Iranian language points to an initial laryngeal (†Hā̆ri̯a- vel sim.): A word-initial laryngeal should have left direct traces in Persianide languages (see Kümmel 2018), but Old Persian <ariy-> /ariya-/ or inscriptional Middle Persian ēr ‘Iranian’ do not preserve such a sound. The hypothetical (East) Scythian would be the only Iranian language to preserve it, and independent evidence for this is entirely lacking. Other etymologies equally rest upon highly questionable ad hoc assumptions on Iranian historical phonology and must accordingly be dismissed (e. g. the etymology of Old Turkic täŋri ‘heaven’ that Beckwith 2022: 195, 203 wants to derive from an East Scythian *tagri through the application of an alleged Scythian syllable contact law of nasalization completely unheard of in the specialist literature and remaining without any reliable parallel; on this word rather cf. Georg 2001).It must therefore be conceded that, while it is a priori likely that Iranian tribes were one factor among others in the ethnolinguistic melting pot of the eastern Eurasian steppe some 2000 years ago (the Sakan languages would be a good starting point for further research in this direction), the evidence adduced by scholars in favour of a dominant role of Iranian groups and their languages in the Xiōng-nú empire so far does not follow the rigorous methodological standards of Historical-Comparative Linguistics and is therefore insufficient to allow for any reliable inferences.Etymological analyses of Xiōng-nú glosses in Chinese sources (collected by Pulleyblank 1962, criticised and reanalysed by Dybo 2007), complemented by the interpretation of the so-called Jié couplet, the only short text preserved in the Xiōng-nú language,2 have led to a more promising alternative hypothesis. This hypothesis acknowledges both the multi-ethnic composition of the Xiōng-nú empire as such and the presence of Indo-European and specifically Iranian languages in Inner Asia at the beginning of the common era, yet adds to the complexity the idea that the native language of the ruling dynasty of the Xiōng-nú empire might have been a Yeniseian one (Ligeti 1950; Pulleyblank 1962; Dul'zon 1966; Dul'zon 1968; Vovin 2000; Vovin 2003; Vovin 2007; Werner 2014; Vovin 2020). Yeniseian languages are usually considered remnants or survivors of the original linguistic diversity of Siberia, historically spoken in retreat areas as the result of several waves of superimposition or displacement by expanding Uralic/Samoyedic, Turkic and Tungusic languages. Therefore, Yeniseian languages are also known as Paleo-Siberian languages.3 Several different Yeniseian languages were spoken in the 18th century AD alongside the middle reaches of the Yenisei River and some of its tributaries, yet this probably reflects a northward migration from a point of departure further south, around the headwaters of the Yenisey, the Ob and the Irtyš rivers (see Dul'zon 1959a; Dul'zon 1959b; Dul'zon 1964; Maloletko 1992; Maloletko 2000; Vajda 2019: 194–95; cf. also Janhunen 2020: 167). From the six historically attested Yeniseian languages Ket, Yugh, Kott, Assan, Arin and Pumpokol, it has so far been suggested that Ket/Yugh (Ligeti 1950; Pulleyblank 1962) or Pumpokol (Vovin 2000, 2003, 2007, 2020; Vovin et al. 2016) may have been the native language of the Xiōng-nú ruling dynasty.Adding value to this hypothesis is the fact that the northward migration of Yeniseian-speaking groups, as reflected in toponyms, from the Altai-Sayan area would well agree with detailed historical studies considering Indic, Iranian and Chinese written sources (de la Vaissière 2005; de la Vaissière 2014). These studies indicate that, following the eventual demise of their steppe empire, remnants of the Xiōng-nú migrated to the north of the Altai-Sayan Mountain ranges in the mid-2nd century AD and that this retreat area was the starting point of a secondary expansion of Xiōng-nú descendants roughly two hundred years later, between ca. 350–370 AD. This expansion occurred in three directions: One migratory trajectory led northward and left traces in the form of toponyms. This population movement downstream of the major rivers Yenisey, Ob and Irtyš perfectly explains the linguistic situation as documented for the first time in the 18th century and provides a direct link between Yeniseian languages and the Xiōng-nú. Another migratory route led to southern Asia and involved groups known from Iranian and Indic sources as Chionites, Kidarites, Hephthalites, Alchons as well as the so-called Huṇa (cf. Pfisterer 2013). A third migratory trajectory led westward, into Europe and involved the Huns who appeared in Eastern Europe in 370 and posed a threat to Roman hegemony until Attila's death in 453, the Battle of Nedao shortly afterwards and the ensuing disintegration of their confederation (cf. e.g. Heather 1996; Bóna 2002; Halsall 2007; Schmauder 2009; Maas 2014; Pohl 2022).Several nomadic groups of late Antiquity that originated in Inner Asia and migrated to the southern and western peripheries of the Eurasian landmass apparently used the same ethnonymic constituent (Chion-ites – Al-chon – Huṇa – Huns; cf. de la Vaissière 2005; de la Vaissière 2014, but see Atwood 2012), and the traditional hypothesis of a link between the ethnic core of the European Huns of the 4th–5th centuries AD and the Inner Asian Xiōng-nú of the 3rd century BC–2nd century AD, first proposed by the French scholar Joseph de Guignes in the 18th century, has, strictly speaking, never been falsified (de la Vaissière 2005: 15).
A genetic connection between the Xiōng-nú and the Huns is usually considered unlikely in modern archaeological and historical scholarship (e.g. Beckwith 2009: 72; Savelyev & Jeong 2020; Pohl 2022; Maenchen-Helfen 1944–1945; Maenchen-Helfen 1955; Maenchen-Helfen 1973; Schmauder 2009), partly because of the large chronological gap between the dissolution of the Xiōng-nú empire in the 2nd century AD and the appearance of the Huns in the 4th century AD, and partly because only two archaeological features render a connection likely: large bronze cauldrons of a certain type and artificially deformed or elongated skulls (Pohl 2022: 147).Despite the prevailing scepticism of historians and archaeologists, the hypothesis of a connection between the Xiōng-nú and the Huns has been corroborated recently by previously unknown and unavailable genetic data analysed by Gnecchi-Ruscone et al. (2025): ‘(…) long-shared genomic tracts provide compelling evidence of genetic lineages directly connecting some individuals of the highest Xiongnu-period elite with 5th to 6th century AD Carpathian Basin individuals, showing that some European Huns descended from them’.
On the provision that there was indeed some continuation between the ethnic core of the European Huns and the former Xiōng-nú, the ruling classes of both multi-ethnic confederations may have spoken the same language in two different diachronic stages (an older form and a younger one), implying that the identification of the linguistic affiliation of one of these groups probably also means identifying the native language of the other group.
In the following, we will discuss previously unknown linguistic evidence from four domains independently supporting such a connection and thus corroborating the recent archaeological and genetic findings: (1) loanwords, (2) glosses, (3) anthroponyms and (4) toponyms/hydronyms.
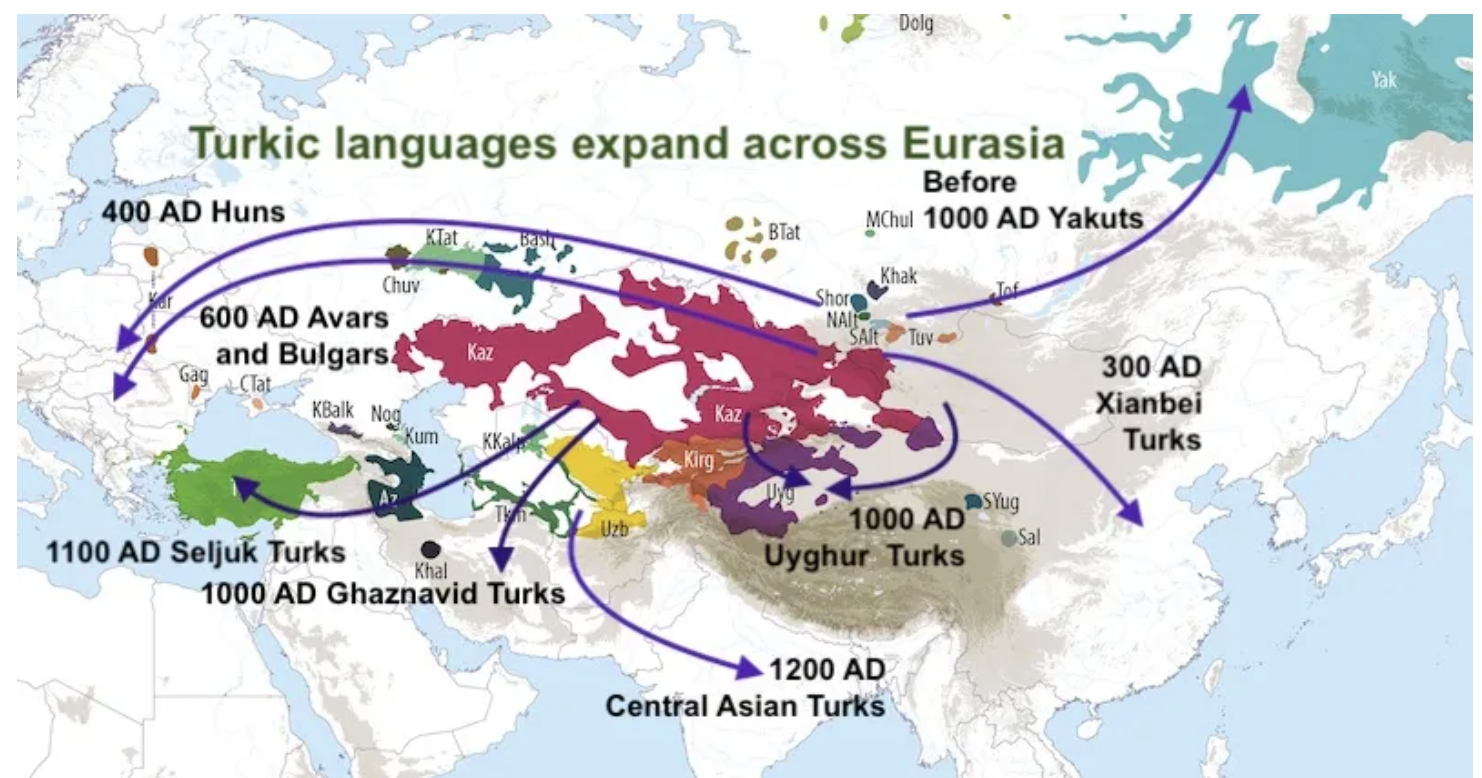
This analysis, which moves the Turkic and Tungistic migrations several centuries later in history than previously believed, is also relevant to the Altaic linguistic hypothesis and our understanding of these ethnic mass migrations more generally.
Close in time and space: Slavic ethnogenesis

The Slavic people emerged around the same time as the fall of the Roman Empire and the demise of the short lived Hunnic Kingdom in the Balkans, but before the Magyar conquest of what is now called Hungary and before the appearance of Gypsies in Europe. This period was traditionally called the "Dark Ages" in Europe. There are some historical roots, however, which suggest Slavic origins several centuries earlier (from the Wikipedia link at the start of this paragraph):
Ancient Roman sources refer to the Early Slavic peoples as "Veneti", who dwelt in a region of central Europe east of the Germanic tribe of Suebi and west of the Iranian Sarmatians in the 1st and 2nd centuries AD, between the upper Vistula and Dnieper rivers. Slavs – called Antes and Sclaveni – first appear in Byzantine records in the early 6th century AD. Byzantine historiographers of the era of the emperor Justinian I (r. 527–565), such as Procopius of Caesarea, Jordanes and Theophylact Simocatta, describe tribes of these names emerging from the area of the Carpathian Mountains, the lower Danube and the Black Sea to invade the Danubian provinces of the Eastern Empire.Jordanes, in his work Getica (written in 551 AD), describes the Veneti as a "populous nation" whose dwellings begin at the sources of the Vistula and occupy "a great expanse of land". He also describes the Veneti as the ancestors of Antes and Slaveni, two early Slavic tribes, who appeared on the Byzantine frontier in the early-6th century.Procopius wrote in 545 that "the Sclaveni and the Antae actually had a single name in the remote past; for they were both called Sporoi in olden times". The name Sporoi derives from Greek σπείρω ("to sow"). He described them as barbarians, who lived under democracy and believed in one god, "the maker of lightning" (Perun), to whom they made sacrifice. They lived in scattered housing and constantly changed settlement. In war, they were mainly foot soldiers with shields, spears, bows, and little armour, which was reserved mainly for chiefs and their inner circle of warriors. Their language is "barbarous" (that is, not Greek), and the two tribes are alike in appearance, being tall and robust, "while their bodies and hair are neither very fair or blond, nor indeed do they incline entirely to the dark type, but they are all slightly ruddy in color. And they live a hard life, giving no heed to bodily comforts..."Jordanes describes the Sclaveni as having swamps and forests for their cities. Another 6th-century source refers to them living among nearly-impenetrable forests, rivers, lakes, and marshes.Menander Protector mentions Daurentius (r. c. 577 – 579) who slew an Avar envoy of Khagan Bayan I for asking the Slavs to accept the suzerainty of the Avars; Daurentius declined and is reported as saying: "Others do not conquer our land, we conquer theirs – so it shall always be for us as long as there are wars and weapons".
The Slavic languages are a relatively recent offshoot of the Indo-European Baltic languages, which in turn may be the most direct descendants of the language(s) of the Corded Ware culture (ca. 3000 BCE to 2350 BCE).
Eurogenes reports on new ancient DNA driven discoveries drawn from the earliest ancient Slavic DNA at his blog.
A paper dealing with the origin of Slavic speakers, titled Ancient DNA connects large-scale migration with the spread of Slavs, was just published at Nature by Gretzinger et al. (see here).The dataset from the paper includes ten fascinating ancient samples from Gródek upon the Bug River in Southeastern Poland. These individuals are dated to the so called Tribal Period (8th – 9th centuries), and, as far as I know, they represent the earliest Slavic speakers in the ancient DNA record.The really interesting thing about these early Slavs is that they already show some Germanic and other Western European-related ancestries. Nine of the samples made it into my G25 analysis (see here). In the Principal Component Analysis (PCA) plots . . . five of them cluster near present-day Ukrainians, while the rest are shifted towards present-day Northwestern and Western Europeans. . . . GRK015, a female belonging to Western European-specific mtDNA haplogroup H1c, shows Scandinavian ancestry. On the other hand, GRK014, a female belonging to the West Asian-specific mtDNA haplogroup U3b, probably has Southern European ancestry.
These results aren't exactly shocking, because the people who preceded the early Slavs in the Gródek region were Scandinavian-like and associated with the Wielbark archeological culture. In other words, they were probably Goths who also had significant contacts with the Roman Empire.However, it's not a given that the ancestors of the Tribal Period Slavs mixed with local Goths. It's also possible that they brought the western admixture, or at least some of it, from the Slavic homeland, wherever that may have been.That's because the early Slavs who migrated deep into what is now Russia also showed Western European-related admixture. This is what Gretzinger et al. say on page 74 of their supplementary info (emphasis is mine):The only deviation from this pattern is observed for ancient samples from the Russian Volga-Oka region, where we measure higher genetic affinity between present-day Southern/Western Europeans and the SP population compared to the pre-SP population (Fig. S17). This agrees with the pattern observed in PCA and ADMIXTURE that, in contrast to the Northwestern Balkan, Eastern Germany, and Poland-Northwestern Ukraine, the arrival of Slavic-associated culture in Northwestern Russia was associated with a shift in PCA space to the West, a decrease of BAL [Baltic] ancestry, and the introduction of Western European ancestries such as CNE [Continental North European] and CWE [Continental Western European].Thus, it's highly plausible that the Tribal Period Slavs from Gródek were very similar, perhaps even practically identical, to the proto-Slavs who lived in the original Slavic homeland. Hopefully we won't have to wait too long to discover whether that's true or not. More Migration period and Slavic period samples from the border regions of Belarus, Poland and Ukraine are needed to sort that out.
Eurogenes goes on to criticize a suggestion in the supplemental materials to the Slavic ancient DNA paper that suggests that
Sycthian groups from Ukraine show varying fractions of South Asian ancestry (between 5% and 12%), a component present in many ancient individuals from Moldova, Ukraine, Western Russia, and the Caucasus, but (nearly) absent in the SP genomes from Central and East-Central Europe (<5%). [Ed. references to specific samples showing this omitted.]
Eurogenes, rightly, explains that the data are really showing European introgression into South Asia arising from the Indo-Aryan invasion of the region in the Bronze Age, and before that from Iran.